第三节吸虫纲(Trematoda)
吸虫纲都是营寄生生活的扁形动物,原始的种类主要寄生在软体动物、变温脊椎动物等的体表或腔隙。也就是主要为外寄生种类,例如单殖吸虫类
(Monogenea)、盾腹吸虫类(Aspidoga-straea)。进化的种类主要寄生在脊椎动物体内,为内寄生种类,例如复殖吸虫类(Digenea)。由于寄生生活方式,吸虫类在形态及生理上产生了相应的适应性特征;例如体表的纤毛及腺细胞退化或消失,出现了附着寄主组织的吸钩及吸盘;消化系统趋于退化, 出现了兼有保护及吸收营养物质的皮膜;内寄生种类行无氧呼吸;神经感官的不发达;强大的繁殖能力及复杂的生活史等都与其寄生生活相关。但吸虫类的幼虫体表仍具有纤毛、并营自由生活,这一点与寄生的绦虫类不同。
一、形态与生理1.外形
吸虫类的身体一般为或长或短的卵圆形、背腹均扁平、前端稍尖、后端稍钝。体长由 1mm—7cm 之间,身体无色或灰色或由于生殖腺及肠道中的食物而使身体呈现黄色、褐色或红色等。
吸虫类显著的体表特征是具有附着器官,在不同的种类其附着器的结构是不同的。原始的单殖吸虫亚纲,主要是用发达的粘着器官附着(图 6-18), 这种粘着器多由腺体构成,或由腺体及肌肉构成的盘,或是体壁凹陷或外突, 其上有粘腺,而且附着器多出现在身体的一端或两端。如为两端则分别被称为前吸器(prohaptor)和后吸器(opisthaptor)。一些种在后吸器的中央还形成 2—4 个角质化的钩(hooks)或锚(anchors),边缘还有许多小钩,
它们用粘液或吸钩附着在寄主体表,例如三代虫(Gyrodactylus)(图 6- 18A),还有的种粘着盘上出现一些凹陷,形成小的吸附器官,例如似多盘吸虫(Polystomoides)(图 6-18B)。凹陷之间也有放射棘相分隔,总之单殖吸虫的吸器上总是伴有钩、刺及角质化的网以起支持作用。在复殖吸虫类, 粘着器官不发达,即使有粘着器也没有钩或锚伴随。它们的附着器官是与体壁分离由肌肉环绕的碗状吸盘,是靠吸盘的真空原理吸着在寄主体内,而不是粘着或钩着。一般具有两个,前端的称口吸盘(or-al sucker),环绕在口的周围;另一个称腹吸盘(ventral sucker),在身体的腹面,例如中华枝睾吸虫(Clonorchis sinensis)(图 6-19)。如果两个吸盘均存在时也可称双盘吸虫( Distome),如果后吸盘次生性的消失则称为单盘吸虫
(monostome)。
吸虫的口通常位于身体的前端,生殖孔位于身体的腹中线上。2.体壁
吸虫的体壁曾一度吸引着许多研究者的兴趣。过去一直认为体壁的最表层是一层角质层(cuticle),由深层的间质细胞分泌形成,是非生活物质。自 1963 年 L.T.Threadgold 用电子显微镜对肝片吸虫(Fasciolahepatica) 的研究,从而否定了用光学显微镜研究所得出的结论,证明它是生活物质, 并与深层的细胞有原生质相联,以后他又用电子显微镜对许多吸虫及其不同发育阶段的幼虫进行研究,结果还证实了它们的体壁中有线粒体
(mitochondria)存在,这就更确定了吸虫的体壁表层是具有代谢活性的生活物质。
吸虫体壁最外表的这层原生质层,没有细胞核及细胞膜,而是一层合胞
体(syncytium)(图 6-20),其中包含有线粒体及内质网(endoplasmic reticulum)、结晶蛋白所形成的刺以及胞饮小泡等,故称这层原生质层为皮层(tegument)。皮层基部为基膜(ba-sement membrane),只是它的细胞体及细胞核均下沉到实质中,由这些细胞伸出细长的管道(trabeculae)穿过肌肉层而与表面的原生质层相联。皮层的这种特殊结构及形态,不仅可以对抗寄主体内消化酶的作用,而且对吸虫通过体表吸收寄主的营养物质及进行气体交换都有重要的作用。
吸虫由于寄生生活,皮层没有纤毛、杆状体等结构。皮层之下为肌肉层, 外层为环肌、内层为纵肌;有的种还有斜肌,皮层与肌肉层也构成皮肌囊。肌肉层之内即为实质,身体两侧的实质中也有背、腹肌穿行。肌细胞亦为平滑肌,吸盘及咽处的肌肉特别发达。
- 营养
吸虫类还保留有消化道,消化系统的结构很简单,包括口、咽、食道及肠,没有肛门。口周围常有吸盘围绕(图 6-19)。咽壁具有肌肉便于抽吸食物、食道很短、肠为单枝或双分枝,其末端为盲端,也称为肠盲囊(intestinal caecum)。
吸虫类均生活在有机质丰富的环境中,特别是内寄生的种类。随着寄生部位的不同,可以取食寄主的上皮细胞、血液、粘液等各种组织及组织的排出物,甚至可取食由寄主直接而来的食物颗粒。寄生在人体肝门静脉中的血吸虫(Schistosoma),直接以寄主的红血细胞为食。用体外培养实验证明: 雄性血吸虫每小时取食 30000 个红血细胞;而雌性成虫是雄虫取食量的 10
倍,特别是在产卵阶段,一头雌虫每日产卵 1000 粒,要消耗掉其自身体重的10%,所以必须要从食物中获取大量的蛋白质及氨某酸。血吸虫取食后先行胞外消化,再行胞内消化。其胃层的上皮细胞形状可由柱形到扁平形,胃层细胞的基部含有非特异性酯酶,在血吸虫取食过程中,酶逐渐由基部移到细胞端部,当酶达到相当数量之后,细胞顶端破裂,酶被释放到肠腔与食物混合进行胞外消化过程;部分的血红蛋白的分解是在细胞内完成。代谢产物及不能被吸收的铁离子经过实质细胞再由排泄器官排出体外。在血吸虫的消化道中也发现了蛋白水解酶,它可以分解球蛋白成为肽的片段。
吸虫类不仅可用消化道取食营养,其体壁的皮层对营养物质的吸收也具有重要的作用。许多溶解于虫体周围环境中的小分子物质可以通过吸虫类的皮层原生质膜的扩散作用(diffusion)从环境中吸收;环境中一些较大的颗粒吸虫类可以通过皮层的主动吸收(吞噬或胞饮)而获得营养。由于皮层内含有来自于自身的内源性酶(intrinsic enzymes)或来源于寄主的外源性酶
(extrinsicenzymes),它可以从周围环境中通过其体壁而吸收营养物质。4.呼吸与排泄
吸虫类与涡虫相似,没有专门的呼吸器官。外寄生的吸虫类以及内寄生种类的营自由生活的幼虫阶段都是行有氧呼吸(aerobicrespiration),即通过体表进行气体交换。而内寄生的种类,由于周围环境中,特别是寄主的消化道内很少有游离的氧,故它们都是行无氧呼吸( anaerobic respiration )。无氧呼吸就是利用贮存在体内的营养物质——糖元
(glycogen)在无氧条件下进行发酵作用以产生能量的过程。这个过程也称为糖酵解作用(glycolysis)。在酵解过程中一分子的葡萄糖借助于酶的作用氧化成两分子的丙酮酸盐,同时释放出两个高能磷酸键,然后再被还原成
二分子的乳酸盐。在有氧条件下,一分子的葡萄糖也被氧化成二分子的丙酮酸盐,但不被还原成乳酸盐,而是经过三羧酸循环将丙酮酸盐进一步氧化成二氧化碳和水,在整个过程中释放出 38 个高能磷酸键。对比这两种异化过程,无氧呼吸是一种不完全的异化过程,它所释放出的能量仅为有氧呼吸的1/19,其代谢的终产物是乳酸盐(lactate)、醋酸盐(acetate)及丙酸盐
(propionate)等中间产物,这是一种低效能的呼吸;而有氧呼吸是葡萄糖的完全氧化,其终产物为二氧化碳及水,并同时释放出更多的能量,比起无氧呼吸有氧呼吸则是高效能的呼吸。
某些内寄生的吸虫,在有氧情况下也可以进行部分的三羧酸循环,但不能完成全过程。也曾有文献报导在一些吸虫的呼吸过程中,有着起重要作用的细胞色素系统,但在无氧呼吸中的意义还是不清楚的。
吸虫类的排泄器官也是原肾,具两条排泄管及大量的焰细胞,单殖吸虫的排泄管分别开口在身体的前端,其代谢产物主要是氨,少数种类也可以产生尿素或尿酸,这些代谢产物在单殖吸虫类也可以通过体表排出,甚至是排泄的主要形式。另外其体内还含有较高浓度的氨基酸,如甘氨酸及脯氨酸等。可能原肾在体内主要是维持渗透平衡作用。在复殖吸虫类的一对排泄管在身体后端中央联合形成一个 Y 形膀胱(bladder),以单个肾孔开口在身体后端中央。在其原肾管液体中曾发现有氨、尿素等,因此具有排泄作用。内寄生种类由于体内、外环境中离子的浓度相对稳定,所以它的原肾对渗透调节作用是很有限的。这在许多内寄生的吸虫中得到证明。
- 淋巴系统
一些复殖吸虫,如同口科 ( Paramphistomidae )及环肠科
(Cyclocoelidae)的一些种,它们的实质中有管系统,由身体两侧的 2—8 个独立的管所组成,这些管分出盲枝分布到肠、生殖器官及吸盘等处,被认为是一种淋巴系统。这些管是由扁平的间质细胞组成,管内有液体,其中也漂浮着游离的细胞,整个淋巴系统似乎担任着食物、排泄物及气体的分散功能。
- 神经与感官
吸虫类的神经结构与涡虫纲相似,包括一对脑神经节(图 6-21A、B), 脑神经节之间有横的神经纤维相联。由脑神经节向前、后分别通出 3 对纵神经索,其中腹神经索更为发达。一般在吸盘及咽处有更多的神经分布。
吸虫类由于寄生环境的稳定性,特别是内寄生的种类,其感官很不发达。感官多存在于生活史中自由生活的幼虫阶段或存在于一些外寄生的成虫。眼点结构也很原始,仅有色素杯,此外在体表散布有感觉毛或感觉乳突,一般在头区较为丰富。感觉器或是由一个神经细胞末端具一根纤毛组成,或是由几个神经末端联合形成。它们担任着触觉、化学感觉及触流感觉等功能,复殖吸虫的感觉毛并不外露,因有体壁的皮层膜所包围。
- 生殖与发育
吸虫类绝大多数种类为雌雄同体,极少数为雌雄异体(如血吸虫)。生殖结构较复杂。雄性系统包括精巢(睾丸)、输精管、贮精囊及生殖孔等(图6-19)。精巢的数目、形状及位置因种而异,常作为分类的依据。例如单殖吸虫亚纲精巢常多个,只有一个输精管;复殖吸虫亚纲精巢多为 2 个、输精管两个,后端联合形成贮精囊,贮精囊常包在阴茎囊(cirrus sac)之中(图6-22A),贮精囊的后端形成射精管(ejaculatory duct)及阴茎(cirrus),
射精管的周围还有前列腺包围,阴茎的末端以雄性生殖孔开口在生殖腔中或单独开口在体外或由生殖腔中外伸出来。
雌性生殖系统包括一个卵巢(图 6-19、图 6-22B),由卵巢通出一条输卵管,输卵管后端接受受精囊管、卵黄腺管,三个管汇合后膨大形成卵膜腔
(ootype),卵在卵膜腔中受精。卵膜腔周围围有梅氏腺(Mehli’sgland), 离开卵膜腔后为一盘旋的管状子宫,子宫末端或开口在生殖腔或直接开口体外,生殖腔有肌肉包围,肌肉的收缩使卵易于排出。许多吸虫输卵管在进入卵膜腔之前,具有一短管,称为劳氏管(Laurer’s canal),其机能不清楚, 可能是退化的阴道,为贮存过多的精子之用。
吸虫在通常情况下行异体受精,有时也发生自体受精。交配时,雄性的前列腺分泌粘液,以保护精子的存活,交配后精子经雌性子宫上游,最后进入受精囊并贮存在其中。卵由卵巢排出后,经输卵管到卵膜腔或在进入卵膜腔之前与从受精囊出来的精子相遇而使卵受精。吸虫的卵也是外卵黄卵,受精卵周围接受卵黄物质,梅氏腺的功能是对卵壳的形成起模板作用或刺激卵黄细胞释放卵黄物质以及活化精子,也有人认为它的分泌物有滑润作用,以利于卵通过子宫。卵经过子宫、生殖孔之后排出体外。与寄生生活相关,吸虫具有巨大的产卵量,与自由生活的涡虫相比,有人估计吸虫的产卵数是涡虫的 10000—100000 倍之多。
吸虫的生活史在不同的种类其复杂程度不同。单殖吸虫亚纲生活史简单,只有一个寄主,由成虫产卵后发育成钩毛蚴虫(on-chomiracidium)(图 6-23),钩毛蚴体表披有纤毛,在水中自由游泳生活一段时间。一般通过化学引诱寻找寄主——鱼,找到寄主后,在附着处如鱼的体表、鳃、鳃腔等部位脱去纤毛,然后变态成成虫。
复殖吸虫生活史复杂,均有寄主更换现象。一般有 2—4 个寄主;两个自由生活的阶段,即寄主转移时期;生活史中有有性生殖及无性生殖时期。在2—4 个寄主中只有一个寄主称终寄主(definitive host),吸虫在终寄主通常是脊椎动物体内行有性生殖。其他的寄主均称中间寄主
(intermediatehost),吸虫在中间寄主体内均行无性生殖,通过无性生殖完成一个或几个世代。中间寄主通常是软体动物的螺类或其他无脊椎动物及植物。换句话说凡行有性生殖的寄主称终寄主,行无性生殖的寄主称中间寄主。
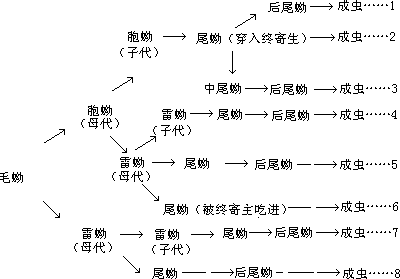
不同种类的复殖吸虫发育过程中所经历的虫态期大致可分为卵(egg)、毛蚴(miracidium)、胞蚴(sporocy-st)、雷蚴(redia)、尾蚴(cercaria)、后尾蚴(metacercaria)及成虫(adult)。但每种的吸虫所经历的幼虫期并不相同,每种幼虫无性繁殖的代数也有区别(图 6-25),这将在复殖亚纲中结合具体虫类来了解。
现将各虫态的一般特征分述于下:
卵吸虫卵大多呈卵圆形(图 6-24A),多数具卵孔,有卵壳包围,由母体产出的卵,多数种在其卵壳内已发育成胚胎或小的毛蚴。卵的大小、形态及卵壳有种的特异性,常用来做鉴别及诊断成体类别的根据。
卵的孵化受外界不同因素影响如水、温度、气压、酸碱度及光照都是影响孵化的因素。例如卵在 10—37℃、pH4.2—9 之外则停止发育,也有的种, 卵是被寄主吞食之后才孵化,这时寄主体内的酶及 CO2 的分压是其孵化的重要因素。
毛蚴毛蚴是生活史中第一个自由游泳的时期,也是感染中间寄主的阶段。身体呈梨形、全身披有纤毛(图 6-24B)。例如日本血吸虫的毛蚴体长仅 80—120μm。身体前端具有顶腺(apicalgland)和头腺(frontal gland), 还具有神经节、眼点、化感器和触觉感受器等,身体两侧有一对原肾,中央有成团的生殖细胞。毛蚴很活跃,在水中自由游泳,以寻找中间寄主。有人认为寄主螺分泌的粘液对毛蚴有化学吸引力,使毛蚴易于找到寄主。也有人认为是由于毛蚴自身的运动使它寻找寄主,毛蚴的年龄决定了它寻找寄主的成功与否,因孵化后 2—5 小时的毛蚴生命力最强,运动范围大易于寻找寄主,超过一定年龄后毛蚴则失去活力,很难找到中间寄主。其他的理化因素, 特别是光也有利于吸引它们寻找中间寄主。当毛蚴发现寄主螺之后,靠头部腺体分泌粘液帮助它穿透表皮组织进入螺体内,并同时脱去纤毛,进入胞蚴期。
胞蚴毛蚴进入螺体后,脱去纤毛成一囊状物,这就是胞蚴(图 6-24C)。除了毛蚴期的体壁、原肾保留之外,其他结构消失。胞蚴多寄生在螺体的足、鳃、触角、淋巴等处,靠体表吸收寄主的营养以供体内胚细胞的生长发育, 一个胞蚴经过无性生殖方式可以产生许多雷蚴或子胞蚴。胞蚴期是吸虫行无性生殖阶段。
雷蚴由胞蚴体内经繁殖出来的小个体即为雷蚴,体长圆形,后端较钝(图6-24D)。具有口、咽及不分枝的肠。雷蚴很活跃,在螺体内爬行,常迁移到螺的肝区及生殖腺处,它们通过肠道或体表吸收寄主的营养。雷蚴也是吸虫类进行无性生殖、扩大种群数量的阶段。由雷蚴经过无性生殖可形成许多尾蚴或子雷蚴。一般雷蚴在螺体内繁殖多代,在螺体内达到相当数量之后才出现尾蚴。一些种类的雷蚴期也可进行寄主的转移。
尾蚴是吸虫生活史中第二个自由生活的阶段,绝大多数吸虫的尾蚴有一长形尾(图 6-24E),有口、口吸盘,肌肉质咽及分枝的肠道。身体前端有穿透腺,其分泌物帮助尾蚴穿透寄主皮肤,也有原肾,其焰细胞的数目及排列是分类及鉴定种属的重要依据之一。尾蚴成熟后,自雷蚴体内出来,并离开中间寄主在水中自由游泳,寻找新的寄主(第二、三中间寄主或终寄主)。故尾蚴是吸虫的第二个寄主转移阶段。尾蚴对光有正趋性,游泳时多漂浮到水面,静止时沉入水中,它在水中的感染力也有一定的时间限制,超过最佳活力阶段,则很难进入新寄主体内。许多实验证明寄主体内的脂肪酸对尾蚴具吸引力。
后尾蚴尾蚴在水中接触到新寄主之后,脱去尾部靠头部的穿透腺进入寄主表皮内、分泌外壁形成胞囊状,称为后尾蚴(或囊蚴)(图 6-24F)。有的吸虫尾蚴进入第三中间寄主(啮齿类),在其体内发育成后尾蚴,例如重翼吸虫(Alaria)。一旦尾蚴或后尾蚴进入终寄主之后或在寄主皮肤上脱去尾部(尾蚴),或在寄主消化道内脱去囊壁(后尾蚴),再经过体腔或肝门静脉移行到寄生部位,如肝、肺、及血液等处变态成成虫。
不是所有的吸虫都经过上述各发育阶段,不同的种有不同的改变,最常见的改变是(1)没有胞蚴或没有雷蚴;(2)多于一代胞蚴或雷蚴;(3)没有后尾蚴,由尾蚴直接感染终寄主,可能的变化举例如下(图 6-25)。