第四节孢子虫纲(Sporozoa)
孢子虫纲全部是营寄生生活的动物,它广泛的寄生于从低等的多细胞动物到脊椎动物各类动物体内。一些种类表现出很强的寄主专一性,即一种孢子虫只能在一种寄主体内生活。生殖方式及生活史相当复杂,都有一个孢子形成期,孢子是其传播阶段。孢子虫类具有很强的繁殖能力。
孢子虫纲一直被认为是亲缘关系很不清楚的一类,但生活史某个阶段的特征对了解其亲缘关系可能提供一些线索,例如一些孢子虫的小配子具有鞭毛,说明它们与鞭毛虫纲有某种亲缘关系;另一些孢子虫能进行变形运动, 说明它们与肉足虫纲有亲缘关系。孢子虫纲所具有的孢子也曾在某些鞭毛虫及肉足纲中出现,因此孢子虫纲可能是异源性起源的一类,对孢子虫纲的分类也带来了困难,后来用电子显微镜的研究发现孢子虫纲中的孢子虫结构并不相同。一些种类的孢子虫及裂殖子具有一个顶复合器(apica1complex); 而另一些种类的孢子虫具有极囊(polar capsule),极囊内有极丝(polar filament)。因此 1970 年 Levine 提出了将孢子虫分为两类;一类为具有顶合器的,称顶合器纲(Apicalcomplexa),也就是我们现在讲的孢子虫纲
(Sporozoa);另一类为具极囊的孢子虫,称为丝孢子虫纲(Cnidospora),
待本章第五节中叙述。一、基本形态及生活史孢子虫纲的动物细胞结构很简单,细胞一般呈圆形或长圆形,细胞质也可分为外质与内质,具有一个细胞核,细胞器不发达,没有伸缩泡、运动细胞器等,或仅在生活史的某个很短时期内出现鞭毛或伪足(例如,疟原虫仅在裂殖子阶段可做有限的变形运动)。孢子虫也没有取食及消化的细胞器,它是通过体表的微孔(micropore) 或细胞膜表面的吞噬作用吸收寄主的营养,属腐生性营养。呼吸及排泄作用通过细胞膜而行扩散或渗透。其孢子虫及裂殖子时期具有顶复合器(图 2- 24)。顶复合器包括顶环(apicalring)、类锥体(conoid)(疟原虫及泰韧虫例外)、棒状体(rhoptry)及微丝(micromemes),这些结构的作用尚不十分清楚,可能与穿刺寄主细胞有关。
孢子虫类具有 1—2 个寄主,其中间寄主可能是蚊、蝇、蛭类及其他动物。终寄主多数是脊椎动物。在寄主体内的寄生部位可能是血细胞、肌肉细胞、体腔、肠道、膀胱及其他体内空间。孢子虫类的生殖方式及生活史相当复杂, 大多数孢子虫的生活史中具有裂殖生殖时期(merogony)、配子生殖时期
( game-togony )及孢子生殖时期 ( sporogony ),称为三段周期
(triphasiccycle)。少数种类生活史中仅有配子生殖及孢子生殖时期,称为二段周期(diphasic cycle)。个别种生活史中仅有裂殖生殖,称为一段周 期 ( phasiccycle ) ( 图 2 - 25 ) 。
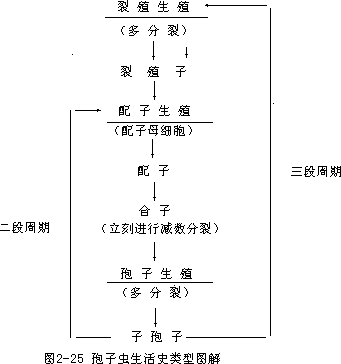
裂殖生殖是指侵入寄主体内的子孢子(sporozoite)所进行的生长之后的多分裂,分裂所形成的许多子体称为裂殖子(merozoite),或称营养子
(trophozoite)。裂殖子是其营养取食阶段,裂殖子先增大体积,再侵入新的寄主细胞,继续进行裂殖生殖,直到寄主体内达到相当多的殖裂子,所以裂殖生殖是孢子虫类的大量繁殖时期。
裂殖子经过多次生殖之后,部分的裂殖子分别分化成大配子母细胞
(macrogametocyte)及小配子母细胞(microgametocyte),由大、小配子母细胞再分化形成大、小配子,大配子不善于活动,也称雌配子
(famalegamete),小配子具鞭毛,可以游动,也称雄配子(malegamete), 然后大、小配子融合形成结合子。所以配子生殖是其有性生殖阶段,结合子是孢子虫生活史中唯一具双倍体(2N)阶段,合子形成后立刻又进行减数分裂,又进入单倍体时期。
孢子生殖是结合子在进行减数分裂之后所进行的分裂生殖,也是单倍体时期。合子形成后能分泌一个很厚的外壁,成为卵囊(oocyst),在卵囊内经过多分裂又形成许多孢子(spore),每个孢子或者不分裂,或者再分裂成2、4 或 8 个子孢子,以后卵囊破裂,子孢子逸出成为传播阶段。所以孢子生殖是在有性生殖之后的无性生殖。
