第四节 生态系统
自从海凯尔(Haeckel)于 1869 年首先创设《生态学》一词后,生物与环境之间相互关系的研究,愈来愈引起人们的重视。在本世纪三十年代,生物学家泰斯莱正式建立生态系统的概念,随后苏联学者苏卡乔夫提出生物地理群落的概念,澳大利亚科学家提出植物环境力学的概念等,其基本实质是一致的,都是关于生物与环境之间能量和物质交换过程的研究。
早在十九世纪,一些学者就表示了这样的意见:原始人类可以灭绝诸如猛犸和其它动物。这种观点受到了在生物进化中反对“灾变论”的人的支持。例如皮多帕里钦科(Pidopli-chko)1963 年就提倡这种看法,他在公开辩论中积极反对只是由于冰期才致某些动物的灭绝。最近(1974 年)布德科也倾向于同意他的观点。如果从生态系统的理论出发,这正是食物链演化中可能发生的过程。
在更新世结束时中纬度动物区系的改变,相当于人类史上旧石器文化时代末期,因而不应当忽视人类在捕杀动物中的作用。某种动物的数目随时间的变化可以表达为:
dn = αn − βn dt
(3.4)
n 为单位面积上某种动物的数目;dn/dt 为该动物数目随时间改变的速率;α为繁殖结果而致的相对增长率;β为由于自然死亡而致的相对衰减率; α及β皆取决于 n 的大小。假如此时人类对于这种动物施加影响时,则
dn = αn − β n − g
(3.5)
dt 1
式中的 g 为给定时间内被人类所猎捕而消灭的动物数;作为自然原因而致的死亡率,β1 在其它条件相同时将比(3.4)式中的β小,因为猎手更容易猎获有病的或体弱的个体。此处当然还应考虑所捕杀动物的性别,在(3.5) 式中只从平均概率出发去描述。
由于所猎获的动物成为当时人类食物的主要来源,则进一步假定 g=γ m,m 为单位面积上的人口数,γ为某给定时间内,每个人所消耗动物物质的数量。现在来考察一下人口的变化,它可以表达成:
dm = am − bm dt
(3.6)
a 与 b 分别表示为人口的出生率及死亡率;我们引入 c=a-b,则可以得出:
m = m ect
(3.7)
mt 为 t 时刻的人口数,m0 为给定时间开始时的人口数。在旧石器时代初期,根据不同作者的推算,在欧洲的人口密度为 5—50 人/100 平方公里,在约 2 万 5 千年当中,应用(3.7)式获得 c 值的范围为 0.64×10-4/年到 1.56
×10-4/年。人类这样一个自然增长率,如与现代人相比,那是十分微小的, 但它却表征了早期人类历史的真实状况,同时也符合由所发现的骨骼去推断当时人类平均寿命的事实。
于是我们进一步估算在欧洲冻原上所存在大量食草动物中的一个品种, 假定它相对于其它动物来说占据统治地位,因而它将消耗这里的初始生产力
(牧草)的绝大部分。但是根据冻原中初始生产力的供应情况去推算,若充
分满足动物食用时,只相当于每平方公里上产生 800 公斤的动物量,此数字决定了这种占绝对优势动物的上限。这个动物如果是猛犸,当其遭到人类捕猎时,其数目变化立即遵循:
dn = (α − β
dt
)n − γm e ct
(3.8)
根据人类增长速率、当地所能产生猛犸的可能数量等推算,猛犸数量随时间的变化直至灭绝,符合于(3.8)式所表达的情况。
从以上看出,对于生态系统中食物链的动态演化问题,很早就已为人们所关注。但只是在近十余年来,生态系统的理论和方法,才逐步完备起来。生态系统具有自己的特点,为了表述它们,将归纳为以下几个方面,现
分别加以简要的叙述。1.生态系统依等级对能量利用的递减性:
在生态系统中,由于各个亚系统所处的地位不同,也就是通常所说的营养级、食物链,其对于能量的摄取与贮存具有递减性的特点。一个保持着正常功能的生态系统,其基础是可直接利用无机环境中的物质和能量,转换成有机的碳水化合物的“初始生产力”。它的水平决定着食草性动物的数量, 而食草性动物的数量又决定着第一食肉性动物与第二食肉性动物的数量。每向上过渡一级,对于能量的总摄取就递降一部分,形成了所谓能量利用的金字塔(图 3.10)。从图上
可以看出,每一级利用前一级生物量中的比例数,决定着本级生物量的多少与生物个体数。可以想象,在同一时刻,一个生态系统中所存留的初始生产力,与其上各级生物中所积贮的能量,它们的总和为一常数,这个常数就是当时当地绿色植物通过光合作用所同化的太阳能数量。表达这种光合作用能力的公式很多,起码有数十种,我们选取:
h 1
b − 1
1440
P0 = 2∫2 ∑ Ana + m n
dt − ∫
Rdt
(3.9)
0 n= 0
I (1 − S)(Tr ) sinπ·t·h 0
此式是蒙泰斯(Monteith,1965 年)、奥斯曼(Osman,1971 年)、申赫(Sheehy,1973 年)等所用公式的修正形式,其中:P0——一日内植物冠丛之净光合作用;A——接受光能的面积;n——表示植物冠丛的层次;其中n=0 和 n=1 分别代表地面和冠丛面;Im——晴朗中午时的光强度;S——冠丛内表示叶子排列方向的参数;Tr——叶子对光的透射率;t——以分钟计的时间;h——冠丛接受光照的时间总数,以分钟为单位;a 及 b 分别是常数;R
——代表植物冠丛的呼吸作用速率,包括光呼吸在内。由于该式考虑了光强度、叶面积、叶子的构造和排列、透射率、光的量子效率(由常数 a 及 b 反映出来)以及植物的分层状况等,因此它是比较全面的。
知道了 P0,也就知道了该时刻生态系统中所贮存的总能量,若以后的各营养级分别以 P1,P2⋯⋯Pn 来表示各级所贮存的能量时,则有
P1 = r0 P0
P = r P = r r P
2 1 1 0 1 0
(3.10)
Pn = rn −1Pn−1 = r0 r1Λ rn−1P0
其中 r0,r1⋯rn-1,分别表示后一级分享前一级能量的百分比。由于 r0,
r1⋯rn-1 均小于 1,故而 r0>r0r1>r0r1r2>⋯>r0r1⋯rn-1,这样营养级越高,其贮存的能量总是越来越小的。同时,各级所贮能量之和:
P0(1-r0)+P1+P2+⋯+Pn=常数=P0
生态系统研究中的一个主要目的,不仅在于确定食物链的组成,而且还需进一步确定 r0,r1⋯的大小,并在系统处于平衡时建立这种 ri 之间的确定比例关系(i=1,2,⋯n)。这种比例的异常,预示着生态系统的变化,这对于捕猎、畜牧、林业、水产等都具有实际的意义。同时,由于 P0 本身决定着整个系统的组成规模,它的大小不仅关乎其上营养级的层次,也关乎这些层次的稳定程度,因此它处于支配作用的地位。我们一再谈到要用它去判定自然地理的质量,用它去衡量自然生产潜力的发挥程度,其原因概出于此。2.物质和能量流动的连续性:
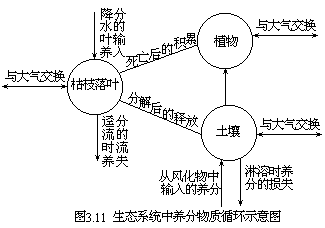
能量与物质一旦输入系统,即连续在各个亚系统之间流动,有时还要进行复杂的反馈,直至最后从系统输出。这种物质和能量的流通过程,既不能中断,也不允许在某个环节阻塞,否则整个系统即行崩溃。例如近来两位地理学家盖斯曼尔(Gelsmehl,1976 年)和敏苔迈耶尔(Meentemeyer,1977 年)在研究了整个生物圈的养分循环后,抽象出了生态系统中极为简化了的养分元素循环图式(图 3.11)。该图明确地表示了物质在系统中的连续流动特性。此外,英国的尤斯海尔(Usher,1972 年),应用莱斯利(Leslie) 在 1945 年提出的著名
矩阵模式,预报了生态系统中能量的交换与磷的循环过程,取得了较好的结果。我们在他的工作基础上,提出了水在生态系统中的连续流动模式, 并且讨论了处理这种连续性的方法,现概略介绍如下:
xi——在任意指定时间内,第 i 个亚系统中水的数量; ai——水输入第 i 个亚系统的流动速率;
zi——水从 i 输出时的流动速率;
fij——水从第 i 个亚系统流向第 j 个亚系统的速率; fii——在一个时间周期内,在第 i 个亚系统中所贮存的水的数量。图
3.12 是水在生态系统中连续流动的模式图。
图中 a1 可理解为对土壤的灌溉;z1 为土壤水分的下渗(此处假定不产生地面迳流),f12 表示水分从土壤向植物的输送速
率;f23 表示植物向大气的蒸腾速率;f13 表示土壤水分向大气的蒸发速率;f32 表示大气降水被植物截留的速率;f31 表示大气向土壤降水的速率。为了加深认识生态系统中物质或能量流动的连续性,就必须揭示出 fij
的物理本质及其数量表达。目前建立各亚系统之间数量关系的可能性已经存在,通过测试和推理即可顺利地求取。现在我们认为系统处于稳定状态,即这种流通速率随时间的变化趋近于 0,此时只要知道 t 时刻各亚系统的值, 通过矩阵处理,可以预测出(t+1)时刻的数值。这无论对于从理论上解释物质或能量在系统中的连续性,还是对于实际上的应用,均是很有意义的。自从莱斯利为处理雌性动物群体的年龄结构提出这一方法后,三十多年来, 得到了在十多个方面的应用和发展。用该法处理上述我们所设计的系统模型,也是完全适用的,在矩阵中出现的 A、B、C 分别是对于 x1,x2,x3 在 t 时刻的净输入速率:
f11 + A
0 f31 x1, t
x1 ,t 1
f f + B f x , t = ,
(3.11) 由于生态系统的各个亚系统间,物
12 22 32 2 2 t 1
f13
f23
f33 + C x3 , t
3,t +1
质能量具有连续传输的特性,这对于我们建立模式、处理模式,提供了十分方便的参考。
3.生态系统发展中的趋稳性:
生态系统的动态演化过程,由于生物与环境间的互相作用与生物对环境的适应性,最后总是趋向于稳定。当其到达稳定之后,只要输入条件不超过稳定态系统的允许范围,则此稳定态得以保持。所谓稳定态,就是说各个亚系统间的物质和能量的流通速率,随时间的变化是 0 或接近于 0。或者说, 亚系统的结构(包括亚系统的组成、层次、分布等⋯⋯)已经定形,没有缺失或增补,物质能量在有限制的输入后(不得过多或过少),在各亚系统间的运转平滑,处于相对平衡状态。
当一种状态达到平衡态时,这时如果一个扰动发生,系统本身有坚持平衡态和抵抗扰动的能力。系统分析中,已经能够将稳定性问题进行数学处理, 并作出判别。
一旦输入条件变化到超越该稳定态的许可边界,则生态系统就由稳定态变成不稳定状态,以后通过生态系统的自我调节及自我适应,必然会在新的条件下达到新的平衡,重新进入一个稳定状态,这就是系统的趋稳性。
植物群落的自然演替过程,充分说明了生态系统趋稳性的特点,它从发生直到演替到它的“顶极”,就是由不稳定到稳定的过程。达到稳定态后, 如果发生了一个强扰动,破坏了这种稳定态,则它又可在新的条件逐步地再达到稳定(见图 3.13)。
