遗传的智慧性——适应性突变之争
获得性状遗传强调外界环境对生物获得新性状的直接作用。虽然确实是有一些生物对环境的变化作出反应并且是可以遗传的例子,但对遗传物质能否受环境的直接作用这一点是不清楚的。而且,大部分的遗传现象还是孟德尔式的遗传,即遗传是以颗粒式基因的稳定性为基础,基因既不发生融合, 也不易发生变化,遗传物质的变异只是偶然的。在这种情况下,一般认为遗传物质的突变或变异是非定向性的,也即是随机的,外界环境的作用最多只能改变遗传物质突变出现的频率,而不能影响突变的方向。事实上,不少突变确是非定向性的。有害的突变出现后,由于影响生物的生存,因此往往被淘汰了;一般的中性突变则随机漂移;而有利的突变即使存在,也是随机产生的。
达尔文主义的理论核心正是认为遗传物质的突变是完全随机的,环境只起到自然选择的作用,突变出现后就要被“适者生存,不适者被淘汰”的原则筛选。因此,达尔文主义是完全否定突变本身可以具有方向性和适应性的。这种观点一直处于统治地位并且是绝大多数人所接受的。
正是由于这种状况,所以在 1988 年,当盖恩斯及其合作者根据他们的发现,在英国的《自然》杂志上发表了一篇题为“突变体的起源”的论文,提出了在环境的作用下,突变的产生可以是非随机的,环境会导致有利的突变以较高的频率出现,从而突变本身就具有适应性的观点后,便在学术界引起了轩然大波。争论不但在生物学家之间进行,还波及到哲学界,并且一直延续至今。
盖恩斯他们的发现是什么呢?在研究最常见的一种细菌——大肠杆菌的变异时,他们设计了一个巧妙的实验。他们选择了大肠杆菌的一个带缺陷的菌株,这个菌株不能利用环境中的乳糖作为“食物”。这是由于这些细菌中与分解乳糖有关的基因有缺陷,因此在一般情况下,必须供给这些细菌其他种类的糖才能使它们正常地生长和繁殖。不过,这些有缺陷的细菌也有可能通过突变而恢复能利用乳糖的功能,虽然产生这种突变的频率一般是很低的。盖恩斯等人用这个带缺陷的菌株作为材料,就是为了研究细菌从不能利用乳糖到恢复利用乳糖的这一突变。他们把这些细菌放入只含有乳糖而不含其他糖的细菌培养基中,由于不能利用乳糖,这些细菌便处于“饥饿”状态, 停止了生长和繁殖。但是过了一段时间后,奇迹出现了,可以看到一些细菌活跃起来,重新开始了生长和繁殖。随着时间的推移,重新活跃起来的细菌不断增多。
有以下几个理由和事实说明这种从不能利用乳糖到可利用乳糖的突变是非随机的,而且是由乳糖诱导产生的:
首先,盖恩斯等人通过对以往证明突变是随机发生的实验进行重新分析,指出以往的实验虽然能证明随机突变确实存在,但是由于所用的选择条件是致死性的,所以只能研究加入选择因素之前出现的突变情况,而不能研究加入选择因素后的突变——已死亡的生物是不会再突变的。因此,以往的实验并不能证明非随机的、由选择因素诱导的突变是不存在的。实际上,很多人都知道这一点,只不过是比较主观地认为所有突变都是随机的。盖恩斯他们所用的选择因素是只提供乳糖,这样即使细菌不能利用乳糖,也不至于马上死亡,而只是处于饥饿状态,并且能继续生存相当长的一段时间。因此,
用这种非致死性的选择条件才能研究加入选择因素之后的突变情况。另一个与以往的实验不同的是,以往只研究生长时期的突变,而盖恩斯等人研究的是在生长停顿期的突变。
第二,在盖恩斯等人的实验中,细菌从不能利用乳糖到能利用乳糖的突变是在被放入只含乳糖的培养基后才产生的,而且随着时间的推移,这种突变不断增多。从而,这种突变是由乳糖诱导的,虽然根据这一点还不能说明只有乳糖才能诱导出这种突变。
第三,把乳糖缺陷型细菌放在不含任何糖的培养基中,使细菌处于饥饿状态,但这样并不能诱导产生出能利用乳糖的突变体。这说明乳糖的存在对产生能利用乳糖的突变体是必需的。
第四,在乳糖诱导下不断产生能利用乳糖的突变体的细菌种群并不能不断产生其他与此无关的突变体,这说明乳糖诱导产生的突变体是有特异性的,即并不是任何突变都能被乳糖诱导产生。
综合上述几点,盖恩斯等人就得出了如下结论:细菌在停止生长的时期存在着一些机制,这些机制使它们能够在环境的作用下,只产生最合适的突变。
这种现象提出后,被冠以多种称呼:如适应性突变、定向性突变、盖恩斯式突变、选择引致的突变、生长停滞期的突变等等,真是五花八门,叫人一下子无所适从。不过,这种现象其实是符合拉马克的进化学说的,即环境对生物有直接的作用,生物在进化中具有对环境变化作出反应的能力。从现代生物学的角度看,适应性突变可以看作是一种遗传的智慧性。适应性突变的观点是属于现代的拉马克主义,因此受到现代达尔文主义的攻击是不足为奇的。
现代达尔文主义者从各个方面试图找出盖恩斯等人的实验的不完善性和漏洞,或对他们所观察到的现象提出另外的解释,希望能在所有突变都是随机的这样一个理论框架中说明这些现象。这种努力导致很多反对适应性突变的文章相继发表。
适应性突变的支持者当然也不甘示弱。他们不断地深入研究,以更严格的手段找出了更多的适应性突变的例子,并且提出了适应性突变的具体机制。这些机制包括在生长停滞期存在着特异的 DNA 代谢(合成与修复)途径, 从而可以导致基因组中一些特异区域的变异;存在着一种对 DNA 的变化进行“尝试”的途径,以只保留有用的变化,使之成为可遗传的变异等等。所提出的机制都是得到一些实验结果的支持的。这样,通过多方面的努力,使得适应性突变现在已经成为一个不容忽视的现象,从而必须认真地考虑其进化意义。
在达尔文主义的进化模式中,生物体的遗传物质可以偶尔随机和无方向性 地 产 生 突 变 , 这 些 突 变
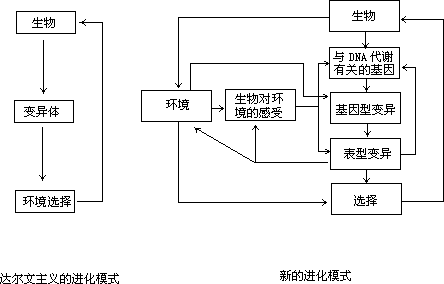
图 12 环境与生物之间相互作用的进化模式通过影响生物体的外表形态或新陈代谢而
表现出来,然后受自然选择的作用,有利的、使生物更适应环境的突变就被保留下来,而很多对生物的生存不利的突变就被淘汰掉。生物进化就发生在“随机突变—自然选择”这样一个不断循环的过程中,环境只是在突变体产生后而被选择的阶段才起作用。而适应性突变的存在就必须要有一个更全面的进化模式才能说明生物进化中的各种情况(图 12)。
在新的更全面的进化模式中,环境与生物体之间可以在多个层次上、以多种方式相互作用,并存在着遗传变异和环境选择之间的反馈效应(图 12)。一方面,环境当然可以对已产生的变异体进行选择,另一方面,生物体通过感受环境的变化,可以改变基因的表达方式,还可以影响遗传物质本身的变异途径,使适应性突变出现。此外,环境中的辐射或某些化学因子也可以直接导致遗传物质的变异。遗传物质的各种变异(又称为基因型变异)导致生物体形态结构或新陈代谢的变异(又称为表型变异),表型变异又可以在一定程度上反作用于环境,引致环境也发生某些变化。在生物体的遗传变异中, 随机性变异受环境的选择或随机漂移,或被保留或被淘汰,而适应性突变都是产生成功的突变体。在这种意义上,遗传物质系统并不总是惰性和被动的, 它们还有智慧性的一面,这无疑对生物适应性的起源和进化有很大的作用。
