小进化与大进化
在生物进化主要是渐进性还是跳跃性这一争论中,不可避免地产生了有关进化的过程是否只有一种的问题。一直以来,有相当多的进化论者认为在物种之内发生的进化过程可能与一类生物进化为另一类生物的过程不同,他们把前一种过程称为小进化或微观进化,而后一种过程就称为大进化或宏观进化。对小进化和大进化到底是不是两种不同的进化过程存在着不少争论, 达尔文主义与非达尔文主义在这一点上是有比较大的分歧的。
小进化与大进化到底是不是一回事呢?
在一个物种内的进化往往都是种群内发生的比较微小的变异,因此小进化其实主要就是物种内的一般变化。实际上,很多物种内的进化是可以观察到的,从而对小进化的研究也就比较容易,还可以从遗传学的角度来对其进行分析。
从遗传学的角度来研究小进化主要就是研究物种内基因频率的改变,这种改变是小进化的一个具体表现。对此,科学家们已进行了比较多的研究。我们知道,一种生物的所有基因在其基因组中都是有各自固定的位置
的,也就是说,这种生物的每一个基因都在其特定的染色体上占有特定的位置。通常,染色体某一特定位置上的基因是有几种不同的形式的,它们就统称为等位基因。在一个双倍体的个体中,最多只能有等位基因的其中两种形式。例如,决定人的 ABO 血型的基因有三种等位基因,而一个人只能有这三种等位基因中的一种或两种(它们决定了一个人的实际血型)。这样,同种的不同个体所携带的等位基因的组合情况就决定了等位基因各种形式的相对多寡。
在一个物种或一个种群中,等位基因的各种形式各自所占的比例就是等位基因的频率,即基因频率(图 35)。基因频率是有相对的稳定性的,不过也有很多因素可以导致整个物种或其中一个种群的基因频率发生改变,从而引起物种内的进化。
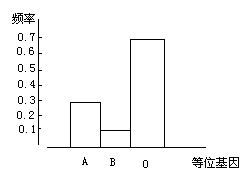
图 35 人类 ABO 血型的基因频率
第一个因素是遗传物质的突变,通过突变有可能会出现新的等位基因, 从而使原来的基因频率发生改变。不过在正常的情况下,突变出现的概率是很小的,即要很长时间或很多个世代才会出现突变,因此一般对短期的种内进化是作用不大的。
第二个因素是自然选择的作用,通过选择使原来的基因频率发生改变。由于在特定的环境下,一些等位基因能使生物更好地生存繁殖,因此它们的比例就会逐渐升高,即这些等位基因在种群中能够扩散。相反,如果是对生
物的生存繁殖不利甚至有害的等位基因,则其比例就会逐渐减少,甚至降低到零,即有关的等位基因被淘汰。对于自然选择可以改变基因频率这一点, 即使是非达尔文主义者也是承认的。
第三个因素是生物个体在不同种群间的迁移,从而改变了有关种群的基因频率。迁移不但可以在动物种群之间发生,也可以在植物种群之间发生。虽然植物不能像动物那样自主运动,但可以通过由动物携带其种子等途径进行迁移。光是迁移这一个因素并不能改变整个物种的基因频率,不过再加上自然选择的作用,结果便有可能是加快或减慢了基因频率的改变。
改变基因频率还有一个因素是遗传漂移或漂变。这是由一些偶然的因素,如有性生殖中的随机交配,而导致的基因频率在后代中的改变。
在上述这几种能够导致基因频率改变的因素中,突变是产生新的等位基因的根本原因;自然选择则是引起基因频率发生变化的强有力因素,在特定的选择条件下,基因频率的改变往往是定向的,即遵循“适者生存,不适者被淘汰”的原则;迁移和遗传漂移也在一定程度上影响着基因频率的变化。实际上,基因频率发生改变往往都是这几种因素中的两种或以上共同作用的结果。
由于一种生物有很多基因,而每一基因都可有多种等位基因形式,从而通过基因频率改变而造成的种内进化是十分多样的。这样,如果一个物种的两个种群被隔离而且生活在有所不同的自然环境中,那么由于基因频率的改变,势必会导致种群的分化。只要这种状况持续的时间足够长,就可以形成不同的亚种。亚种是物种以下的分类单位。和物种一样,亚种也是有特定的地理分布区域的,不过其分布区域往往只是整个物种分布区域的一部分。一个物种的亚种之间在形态学与生理学等方面存在着可以遗传的差异,但它们仍然可以相互交配并产生可育的后代,因此是属于同一个物种。
实际上可以观察到不少亚种快速形成的例子。麻雀在 19 世纪中期才被人
从欧洲带到北美洲。20 世纪 30 年代,有一种群迁移到墨西哥,到了 60 年代便在那里产生了一个亚种,其羽毛的颜色和喙的大小与其他地区的麻雀有明显的差异。这是对研究种内进化很有意义的一个例子。
由于对小进化,特别是对种内基因频率变化的研究取得了不少实质性的进展,从而使得很多进化生物学家,特别是研究遗传学的进化生物学家,一度认为生物进化就是基因频率的变化。他们认为,所有生物进化的过程,包括新物种的形成,甚至还包括新类型生物的产生,都是与小进化的过程相同的。但是,这样的一个推论却是没有什么事实依据的。
在漫长的进化历史中,大多数进化现象都是与生物体内的复杂结构、器官系统、以至整个个体的重大变化有关的。通过基因频率的变化能够说明物种内的小进化,但这对于说明有重大变化的宏观进化却是无甚帮助的。没有任何证据表明大进化现象同样是由引起小进化的遗传机制引起的。
大进化现象虽然普遍存在,但其具体过程却几乎无从考据,因此只能通过推断而得知。很多达尔文主义者都认为大进化只不过是物种内进化的放大,而这一论点遭到了要同时研究不同类型生物的形态学家和古生物学家的普遍反对。非达尔文主义者也认为大进化与小进化是两种不同的过程。
实际上,大进化一直是通过比较生物的外部形态及其功能来加以研究的。虽然现在还不清楚在大进化中遗传物质到底发生了什么变化,但已经基本上可以肯定这种变化是不能还原成小进化中的遗传变异的。
生物种类虽然千千万万,但它们可以归纳为数目不多的一些大类,如脊椎动物、昆虫、被子植物等。每一大类都有很多共同的特点,如近 100 万种
的昆虫都有 3 对足,哺乳动物的基本结构特点在大象、蝙蝠、鲸、人类中都是一样的。而另一方面,大类之间却似乎有难以逾越的鸿沟。如何从一大类型的生物进化到另一大类型有较近亲缘关系的生物,例如从爬行动物进化到鸟类,是生物进化研究中的一大难题。无脊椎动物现存的大部分门类都是在进化的较早时期并在较短的时间内产生的,此后就一直没有什么重大变化。古生物学中对化石生物的研究也发现,新的种类是在较短时间内形成的,然后就有一段很长的进化停滞时期。所有这些现象的存在说明不能光以研究小进化的方法来研究大进化。小进化与大进化是生物进化中的两个不同的层次,把它们区别对待是有利于进一步深入研究生物进化的问题的。
