二、生物不同层次的结构
什么是生物?我们可以说生物就是活的物体,或者说有生命的物体。生物包括动物、植物和微生物三大类。一切活的生物表现出来的共同特征是, 它们都能进行新陈代谢、生长、发育、繁殖、遗传、变异、适应和对刺激的反应等。所有这些特征都可以认为是生命运动的具体反映。
- 生命的物质基础
生命是一种物质的高级运动形式,这种运动形式本质上就在于不断进行以核酸和蛋白质为主体的生物大分子系统的自我更新。组成生物体的分子完全遵守所知的化学规律。在生命运动中这些分子的相互作用固然有其特殊的法则,然而这些法则与物理世界所发现的规律和作用力是相一致的,并不存在什么生命驱动力。
然而,生命运动又是一种特殊形式的运动,有别于非生命物质运动的形式。生命运动的特点在于它的自主性、连续性和适应性。生命的自主性表现在生命系统的新陈代谢是自我完成的。它不断从外界环境摄取物质和能量, 用以构造自己,这是同化作用;同时又不断分解自身的成分,释放能量,并将分解产物排出体外,这是异化作用。由一系列同化作用和异化作用形成的复杂过程就叫做新陈代谢。这种自我完成的新陈代谢,即自我更新,一旦停止,生命也就随之结束。生命的连续性在于生命系统能通过新陈代谢进行自我繁殖,从而繁衍后代。繁殖具有特异性,子代总是与其亲代相似,这就是遗传。生命的适应性在于生命系统具有自动控制的能力,它能随机应变地适应周围的环境。
现代生物学的研究表明,一切生命现象都是跟核酸、蛋白质等生物大分子分不开的,生物大分子是生命现象的物质基础。生物大分子的自复制、自组装和自调节决定了生命的自主性、连续性和适应性。核酸分子能够自我复制。由核酸控制蛋白质分子的合成,并通过蛋白质合成其他生物分子。生物大分子具有自我组装的能力,即由生物大分子聚合成为超分子结构,进而形成细胞。细胞的生长和分裂是在生物大分子复制和合成的基础上进行的。细胞通过分化而形成多细胞生物的组织、器官和个体。这就是说,生物不同层次的结构是生物大分子自组装的结果。再者,生物大分子的功能具有自我调节的作用,从而能够适应细胞内外环境条件的变化。
自然界任何系统的运动总是包括物质、能量和信息三个方面。生命系统的新陈代谢也包括物质代谢、能量代谢和信息代谢三个方面。物质和能量的概念是大家所熟悉的。那么什么是信息呢?简言之,信息表示系统的组织结构,它可以作为系统组织程度(即有序性)的量度。我们通常所说的生物信息或遗传信息主要就是指核酸和蛋白质分子的结构信息。常识告诉我们,从无序状态进入比较有序的状态,总是需要投入能量,这一过程决不会自发地出现。相反,从有序变为无序却是自发而无需提供能量的过程。生物体在形成复杂的组织结构时,必须摄取能量,并且破坏周围和环境的有序结构,也就是说通过增加环境的无序性来形成局部的有序结构,这就是生命系统的信息代谢。
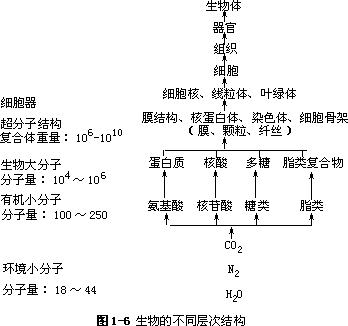
生物功能是由生物结构所决定的,由亲代传递给子代的生物信息控制着子代的结构形成。生物具有不同层次的结构(图 1-6),在此基础上进行各种生命活动。我们对生命世界的认识也是从生物结构开始的。
- 生物大分子
在分子水平上生物体可看成是一个复杂的化学系统,这个系统包含进行新陈代谢、生长、发育和繁殖所必需的全部信息。现有生物体在化学成分和结构方面都有共同的特征,表明它们有统一或类似的起源。组成生物体的分子,除水和无机盐外,各种有机成分都是由生物所合成,因此这些有机化合物都被称为生物分子。水是生物体内最大量存在的分子,它参与一切生命活动,维持着生物体的正常功能。水对于生命来说是极端重要的,由此推测, 生命最初可能起源于水中。某些无机离子对生命活动也是不可缺少的。生物体内主要的有机小分子约有 30 多种,包括氨基酸、核苷酸、糖类和脂类等, 它们是合成生物大分子的前体,并广泛参与体内的各种代谢过程。有些有机小分子还具有化学通信作用。
生物大分子共有四类,即蛋白质、核酸、多糖和脂类复合物。前三类分别由氨基酸、核苷酸和单糖聚合而成,就如同一条项链,每个珠子相当于一个小分子单位,连在一起构成一条大分子链。对于某些多糖来说,它们形成树状的分支链。脂类分子仍属于有机小分子,但它们能通过较弱的次级键形成复合物,在生物膜中就是这样,其性质与生物大分子相似。生物大分子所以能成为生命的负荷者,与其独特的性质有关。这些独特的性质是:结构特异性、反应多样性和构象易变性。组成蛋白质的氨基酸有 20 种,蛋白质分子
有大有小,一个中等大小由 300 个氨基酸组成的蛋白质分子,其排列方式为20300,这是一个天文数字。哺乳类动物基因组大约含有十万个编码蛋白质的基因,其 DNA 分子的信息含量远远大于蛋白质。在宇宙中,还未发现信息含量可与蛋白质和核酸相比拟的分子。其次,蛋白质含有众多的化学基团,它还能与各种有机分子和金属离子相结合,以增加特殊的反应基因,这就使蛋白质能参与多种多样的化学反应。在这方面与自然界任何分子相比,蛋白质也是独占鳌头。第三,生物大分子的构象,即其分子通过折叠而表现出来的
空间结构,易受各种因素的影响而发生改变,因而能调节生物大分子的功能。多糖和脂类复合物通常仅由少数单体按一定规则组成,信息含量远比蛋白质和核酸为低,其反应基团也有限,故通常仅和蛋白质分子结合在一起,协助完成某些生命功能。
下面分别介绍各类生物大分子。
- 蛋白质 蛋白质的英文名词是 protein 。该词最初从希腊文“proteios”衍生而来,意思是首位的。在未发现核酸前,人们认为蛋白质是生物体最重要的成分。但后来证明核酸是更为基本的生命物质,因为蛋白质的合成和结构都取决于核酸。然而无疑,蛋白质在生命系统中占据着极为重要的地位,蛋白质参与一切生命活动,生物的性状都要通过蛋白质来表达。如果说生命蓝图掌握在核酸手中,那么多才多艺的蛋白质则是生命功能的执行者。正因为如此,在生物技术工业产品中主要是各种蛋白质。
组成蛋白质分子的基本结构单位是α-氨基酸。氨基酸是含有氨基和羧基的有机化合物,其分子结构式如图 1-7 所示。用于合成蛋白质的氨基酸共有
20 种,它们之间的差别仅在于和α-碳原子(碳链骨架的第一个碳原子)相连的“侧链”(在图 1-7 中用 R 表示)不同。根据侧链的性质,可将氨基酸分为中性、酸性和碱性的,或分为疏水性(非极性)和亲水性(极性)的。

由一个氨基酸的羧基与另一个氨基酸的α-氨基脱水形成的共价键称为肽键(图 1-8),产物为二肽。当许多氨基酸以这种方式连接在一起时,产生的氨基酸聚合物称为多肽,多肽链中氨基酸称为残基。蛋白质是由一条或一条以上非常长的多肽链所组成。在一条多肽链中,一端总是有一个游离的氨基,称为氨基端(N 端);另一端总是有一个游离的羧基,称为羧基端(C 端)。蛋白质或多肽链的化学性质和空间结构主要是由氨基酸侧链所决定。
图 1-8 肽键的结构
生物界存在蛋白质的种类估计在 1010~1012 数量级。造成种类如此众多的原因主要是 20 种参与蛋白质组成的氨基酸在肽链中排列顺序不同引起的。蛋白质的这种顺序异构现象是蛋白质功能多样性和种属特异性的结构基础。从外形上讲,可将蛋白质分为球状蛋白质和纤维状蛋白质两大类。球状蛋白质溶解度较好,能结晶;纤维状蛋白质又可分为可溶性和不溶性两类。不溶性纤维蛋白质在体内主要作为结构成分存在。每一种天然的蛋白质都有自己特有的空间结构,这种空间结构通常称为蛋白质的构象。
为了表示蛋白质结构的不同组织层次,习惯采用下列专门术语:一级结构是指多肽链的氨基酸排列顺序;二级结构是指多肽链主链借助氢键排列成有规律的周期性构象,主要有α-螺旋、β -折叠和β-转角三种基本形式,氨基酸侧链不参与二级结构,但对其稳定性有影响;三级结构是指多肽链借助各种次级键(非共价键)盘曲折叠成具有特定走向的紧密构象;四级结构是
指寡聚蛋白质中各亚基(亚单位)的空间关系,亚基通常由一条多肽链或以共价键(如二硫键)连在一起的几条多肽链组成。
蛋白质的一级结构是由共价键联结的,因此比较稳定。然而,维持蛋白质空间构象的作用力主要是一些弱相互作用或称为次级键(非共价键),包括氢键、范德华力、疏水相互作用和离子键。这些弱相互作用也是维持核酸构象、生物膜结构的作用力,因此被称为是“维持生命系统结构的作用力”。天然蛋白质的构象取决于上述作用的综合结果。图 1-9 表示卵溶菌酶的三级结构。从图中可以看到多肽主链的α-螺旋、β -折叠和β-转角,相邻二级结构(β-折叠)形成的组合体,在此基础上多肽链折叠成近乎球状的紧密构象。
图 1-9 卵溶菌酶的三级结构
天然蛋白质在某些物理因素(如热、紫外线和表面张力等)和化学因素
(如有机溶剂、尿素、胍、酸、碱和重金属离子等)作用下,引起生物活性丧失、溶解度下降和一系列物理化学性质的改变,这种过程称为变性作用。蛋白质变性的实质是分子中次级键被破坏,引起天然构象解体,肽链变得伸展和松散,内部的疏水基团随之暴露出来。当变性因素除去后,有时变性蛋白又能恢复天然构象,这一现象称为复性作用。蛋白质易于变性是有些蛋白质至今未能开发利用的主要原因。因此,提高蛋白质的稳定性是生物工程的一项重要任务。此外,找出变性蛋白质的复性条件,在生产上也有很大意义。
- 核酸 核酸(nucleic acid)最初从细胞核中分离出来,故得此名。后来知道,无论细胞核还是细胞质,或是无核结构的细菌,都存在核酸。核酸可分为两类,即脱氧核糖核酸(DNA)和核糖核酸(RNA)。它们都是高聚物,其构造单位称为核苷酸。核苷酸本身由三个更小的单位组成:一个五碳糖(核糖或α-脱氧核糖),一个磷酸基团和一个含氮有机碱。含氮有机碱主要有五种:腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、胸腺嘧啶(T)和尿嘧啶(U)。DNA 与 RNA 的差别在于前者由脱氧核糖核苷酸聚合而成,后者由核糖核苷酸聚合而成。两者在碱基组成上也有不同,DNA 中含有 A、G、C、T, RNA 不含 T,而代之以 U。
1953 年沃森(Watson)和克里克(Crick)提出 DNA 分子双螺旋结构模型,在分子水平上解释了遗传信息传递机制,从而奠定了分子生物学的基础。按照沃森-克里克模型,DNA 是由两条反向平行的多核苷酸链围绕同一中心轴构成双螺旋的结构。两条链都是右手螺旋,依靠彼此碱基之间形成的氢键而联系在一起。根据分子模型计算,一条链上的嘌呤碱必须与另一条链上的嘧啶碱相配,其距离才正好与双螺旋的直径吻合。而且,为使碱基对之间能够形成氢键,A 只能与 T 配对,可形成两个氢键;G 与 C 配对,形成三个氢键。上述碱基之间配对的规律称为碱基互补。根据碱基互补原则,当一条多核苷酸链的序列被确定之后,即可决定另一条互补链的序列。碱基互补原则具有极重要的生物学意义,核酸的复制、转录和反转录等过程都是基于碱基互补。沃森-克里克的模型为随后各实验室的大量工作所证实,只在某些细节上有所修正。为表彰他们的杰出贡献,1962 年他们被授予诺贝尔奖。
生物体的遗传信息以密码的形式编码在 DNA 分子上,表现为特定的核苷酸序列。DNA 分子的一级结构即是指其核苷酸序列。为要了解基因 DNA 的编码信息就要测定 DNA 的序列。美国科学家准备用 15 年时间,30 亿美元经费,
绘制出人类基因组(由 30 亿个核苷酸对所组成)全部图谱。这是生物科学有
史以来最伟大的一项工程,这项工程已于 1988 年开始。通过这项工程,人类将能更好了解自己,并有助于遗传疾病和肿瘤的诊断与基因治疗。
DNA 的二级结构是指其双螺旋构象。在生物体内 DNA 的两条互补链通常以接近沃森-克里克模型描述的双螺旋结构形式存在,但局部也能形成相反方向的左手螺旋(称为 Z 型结构)。
细菌的染色体 DNA、质粒(即染色体外基因)DNA、某些病毒 DNA、以及真核生物细胞器(如叶绿体和线粒体)DNA 都是环状分子。真核生物的染色体 DNA 是线状分子。当环状 DNA 两条连续的链形成双螺旋时,拧得过紧或过松都会产生超螺旋结构,这就是 DNA 的三级结构。染色体中的 DNA 能和蛋白质结合形成更高级的结构。DNA 的二级、三级和更高级结构在分子识别和功能的控制与调节中起重要作用。DNA 分子的各级结构如图 1-10。
图 1-10 DNA 的分子结构
在变性因素(如热、酸、碱和变性剂等)作用下,DNA 双螺旋结构被破坏,两条互补链解开,此过程称为变性。除去变性因素后,两条互补链又能重新结合,恢复双螺旋结构,此过程称为复性。如果将两种序列相近的 DNA 分子混合在一起,经变性和复性,不仅同种 DNA 两条互补链能重新结合,异种 DNA 两条部分互补链也能通过形成局部双螺旋区而结合在一起,成为异源双链或杂交分子。一条 DNA 链还能和另一条互补或部分互补的 RNA 链形成杂交分子。在生物技术中经常利用分子杂交来鉴定和分离核酸。从基因库中钓出一个基因就是分子杂交技术的一种运用。
RNA 通常是一条单链分子,它的一级结构即是其核苷酸序列。但 RNA 能通过自身回折形成局部碱基配对的双螺旋区,这是 RNA 的二级结构。碱基配对的原则与 DNA 相似,但以 U 取代 T,即 A 与 U 配对,形成两个氢键;G 与C 配对,形成三个氢键。不配对的碱基被排除在双螺旋结构之外,而形成突环。图 1-11 表示 RNA 的一级结构和二级结构。
图 1-11 RNA 的分子结构(a)RNA 链一个小片段的序列
(b)RNA 通过自身回折形成局部碱基配对的双螺旋区
RNA 的种类很多,主要有:核蛋白体 RNA(rRNA)、转移 RNA(tRNA)和信使 RNA(mRNA)。核蛋白体是蛋白质合成场所,由少数几种 rRNA 和几十种蛋白质分子组成。rRNA 含量很大,约占细胞总 RNA 的 80%左右。tRNA 在蛋白质合成中起运载氨基酸和翻译的作用,约占细胞总 RNA 的15%左右。mRNA 是蛋白质合成的模板,约占细胞总 RNA 的 3~5 %,数量虽少,种类却非常多,大小不一。细胞内还有一些分子量较小的 RNA,在基因表达的不同水平上起调节和控制作用。此外,许多病毒也含有 RNA。
克拉克于 1958 年提出中心法则,认为在基因表达过程中遗传信息流是单向的,从 DNA 转录成 RNA,再由 RNA 翻译成蛋白质,信息进入蛋白质后不再转移。DNA 和 RNA 都采用核苷酸来编码信息,信息由 DNA 转移到 RNA,如同磁带倒录,故称为转录;RNA 和蛋白质的语言不同,前者用核苷酸语言,后者
用氨基酸语言,故前者至后者需经过翻译。中心法则可能是争议最多的一条法则了。有趣的是人们怀疑它,可又普遍接受它;想推翻它,却又推而不翻。70 年代初发现致癌 RNA 病毒含有逆转录酶,能以 RNA 为模板合成 DNA。艾滋病病毒也是一种逆转录病毒。逆转录过程的存在说明遗传信息能由 RNA 传递给 DNA。然而蛋白质多肽链合成后立即卷曲起来,它的氨基酸序列从未发现能指导其他生物大分子的合成。看来,对中心法则实质的了解还有赖于生物科学三个最基本理论问题——生命起源、生命进化和生命本质的进一步深入研究。
50 年代以来,因从事核酸或有关领域的研究而获得诺贝尔奖的科学家已有四十多人,这在其他科学领域内是罕见的。这种情况也从一个侧面说明核酸研究发展的迅速和科学界对核酸研究成果的重视。
-
多糖多糖是由单糖聚合而成,单糖之间以糖苷键相连。多糖可以由一种单糖组成,例如淀粉、糖原(动物淀粉)和纤维素都是葡萄糖的聚合物, 淀粉和纤维素的差别仅在于糖苷键的不同,前者为α型,后者为β型。它们主要作为贮能形式(如淀粉),以及结构成分(如纤维素)而存在。杂多糖含有多种单糖和单糖衍生物,并可和脂类分子结合形成糖脂,与蛋白质结合形成糖蛋白、蛋白聚糖等。它们具有各种不同的功能,在保护、免疫和分子识别中起重要作用。多糖与蛋白质和核酸的主要差别在于,它的合成是由酶控制的,无需生物模板,因此它有种族特异性,但无高度复杂的结构信息。此外,多糖常为分子大小不均一的组分所组成。
-
脂类复合物脂类包括的范围很广,所有不溶于水而易溶于有机溶剂的生物分子都属于脂类,于是它们构成了细胞的疏水成分。最常见的脂类物质为油脂,是由脂肪酸和甘油所组成,它们是生物体所需燃料的贮存形式和运输形式。组成生物膜的脂类复合物主要有三类:磷脂、胆固醇(或称为胆甾类)和糖脂。这三类都是两性化合物,既含有亲水的极性“头部”,又含有疏水的非极性“尾部”。这种两性性质在形成生物膜的脂类复合物中起重要作用,在水环境中亲水头部趋向于水,疏水尾部都有不与水接触的强烈倾向, 从而使分子挤在一起。
- 生物膜
生物的基本结构和功能单位是细胞,任何细胞表面都有一层膜(厚度约4~7 纳米)将其内部成分与环境分开,这层膜称为细胞膜或质膜。此外,大多数细胞中还有许多内膜系统,组成具有各种特定功能的亚细胞结构和细胞器。生物膜对细胞的有序反应和整个细胞的区域化控制提供了必要的结构基础,从而使细胞的物质运输、能量转换、信息传递等生命过程得以有条不紊、协调一致地进行。膜结构既是细胞结构的基本形式,也是生命活动的主要结构基础。
- 生物膜的分子结构化学分析表明,生物膜主要由蛋白质、脂类、糖类, 还有水和金属离子等组成。膜的成分随膜的种类不同而有很大差别。
膜蛋白以其功能大致可分为五类:酶类、运输蛋白、运动蛋白、信息接受与传递蛋白、支持与保护蛋白。这些蛋白质赋于膜各种特殊功能。膜脂主要是磷脂,此外还有胆固醇(细菌不含胆固醇)和糖脂。膜的性质和结构与膜脂的两性性质有关。通常膜脂具有一个亲水的极性基团(头部)和两条疏水的碳氢链(尾部)。在水环境中,极性头部与水接触,疏水的碳氢链被水排斥而挤在一起,使膜脂自发形成平行排列的双分子层,并自我封闭成微囊。
除此,脂类分子还能聚集在一起形成微团结构(图 1-12)。生物膜中脂类分子通常总是以脂双层结构存在,或者组成平面膜,或者形成微囊。膜脂具有流动性。脂类分子碳氢链越短,不饱和键越多,流动性也越大。在生理条件下磷脂大多呈液晶态,即排列整齐的液态晶体,当温度降低至一定值时,磷脂从流动的液晶态转变为类似固态晶体的凝胶态。温度升高又能促使凝胶态“熔解”为液晶态。此转变温度称为相变温度。生物膜中常含有两种或两种以上的脂类分子,它们的相变温度各不相同,在一定温度下有的处于凝胶态, 有的处于液晶态,处于凝胶态和液晶态的脂类分子各自汇集,从而使生物膜的不同区域出现不同的状态(相),这一现象称为分相。由此可见膜脂双层结构具有自组装、自封闭、自分相的作用。
图 1-12 膜脂双层结构和微团结构
{ewr MVIMAGE,MVIMAGE, !70000046_0023.bmp}
生物膜中含有寡糖或多糖,它们大多与膜蛋白结合,少量与膜脂结合。膜的糖蛋白可能与细胞表面行为有关,细胞与周围环境的相互作用都涉及到糖蛋白,伸出细胞表面的多糖如同天线搜集外界信息,在细胞识别中起重要作用。
- 生物膜的透性与物质运输 细胞质膜和细胞器膜的通透性是活细胞维持其内环境的相对恒定和进行正常物质代谢的根本保证。现已知道,生物膜的结构状态和膜中存在的许多运输蛋白与膜的选择性透性有关。正如物质代谢是能量代谢和信息代谢的基础一样,膜的能量转换和信息传递均有赖于膜的选择透性造成的膜内外离子浓度差和膜电位差,在此基础上形成膜的许多其他功能。
从能量的角度来看,膜对分子和离子的透性或者说膜的物质运输可分为两大类:被动运输和主动运输(图 1-13)。物质分子透过膜,如果是从高浓度一侧转移到低浓度一侧,此过程无需提供外来能量,称之为被动运输。反之,逆浓度梯度的运输则是一个消耗代谢能的过程,称之为主动运输。
物质的被动运输是一个依赖浓度梯度的扩散过程。非极性分子和小的极性分子能够直接透过膜的脂双层,其通透能力决定于分子大小和脂溶性;但是离子和稍大一些的极性分子则受到膜脂层的阻挡,它们穿过膜要靠运输蛋白。膜上由通道蛋白形成的亲水通道,允许适当大小和带有一定电荷的溶质, 通过简单扩散的方式进出细胞。膜的载体蛋白在物质运输中起重要作用。它可与被运输的物质分子可逆结合,由浓度高的一侧携带分子,然后转向浓度低的一侧,释放出被运输物质,其作用有如旋转门。这种被动运输方式称为促进扩散。载体与被运输物质的结合是高度特异的,正如酶对反应底物具有特异性一样,因此也可将这些载体蛋白叫做渗透酶。
物质的主动运输是一个逆浓度梯度的过程,这种过程必须由某些产能反应来驱动。将产能反应与需能反应相耦联,这是生物用来推动自然界不能自发进行的各种过程的基本原理。在这里,产能反应提供的能量被用来改变载体蛋白的构象,从而使载体蛋白能够来回将物质从低浓度区搬运到高浓度区,能量的耦联是由载体蛋白来完成的。主动运输与被动运输中载体蛋白的差别仅在于是否与能量耦联。根据能量来源,主动运输可分为:①和另一物质运输过程耦联;②和化学反应耦联;③和其他形式能量(如光能)耦联。
图 1-13 生物膜的被动运输和主动运输
动物细胞外部 Na+浓度总是高于内部,这种浓度差可被细胞利用来主动摄取糖和氨基酸等营养成分。载体蛋白在运载一个有机溶质分子的同时也运载了一个 Na+。这样,Na+由于浓度落差释放的能量正好用于溶质逆浓度运输的需要。这种情况与人类利用水力的原理十分类似,人们可以利用水位落差的能量来作各种功,包括将重物从低处提到高处。对于细菌来说,通常与主动运输相耦联的浓度降落物质不是 Na+,而是 H+。
很多离子,特别是 Na+、K+和 Ca+的主动运输,都是和 ATP 的水解相耦联。ATP 即三磷酸腺苷,是细胞代谢通用的能量载体。它含有两个高能磷酸键, 在水解时能释放出大量能量。细胞获得的能量通常以合成 ATP 的方式将其贮存起来,以备需要时利用。动物细胞的质膜上分布着大量 Na+-K+运载体蛋白, 每一运载过程分解一个 ATP 分子,将 3 个 Na+排出细胞外,同时吸收一个 K+ 进入细胞。这种运载体的作用有如泵,故称之为离子泵。泵的工作需要分解ATP,因此具有 ATP 酶活性。Na+-K+泵将 Na+排出细胞外,将 K+吸收进细胞内, 但是 Na+和 K+又能经离子通道渗漏回去。为维持细胞内外 Na+和 K+恒定的浓度差,需要 Na+-K+泵昼夜不停地工作,静止时动物所消耗的 ATP 约有三分之一用于 Na+-K+泵的活动。由此也可以看出,Na+-K+泵在动物细胞生命活动中占有重要的地位。
另一种主动运输过程是与基团转移反应相偶联。例如,细菌在吸收葡萄糖时,载体蛋白将葡萄糖带至质膜内侧即通过基团转移反应使其磷酸化,被磷酸化的葡萄糖不再能透过质膜,从而使葡萄糖得以不断吸收进细胞内。磷酸化需要能量,所消耗的能量用来促进葡萄糖的主动运输。
- 生物膜的能量转换生物体内重要的能量转换过程都与膜结构密切相关。线粒体是进行氧化磷酸化的主要场所,因而有“细胞动力站”之称;植物的光合作用是在叶绿体的类囊体膜上进行的,可以说整个生物界是由它们养活的。细菌的氧化磷酸化过程在质膜或其内陷的膜结构中进行。有些细菌也能进行光合作用,它们的质膜含有菌紫质(细菌视紫红质),能将光能转变为化学能。
线粒体的数目随不同细胞而异,其外形或为线状,或为颗粒状,故有此名。线粒体的内膜有许多向内褶叠的嵴,嵴的存在有利于增加内膜的面积。内膜包围着呈胶状的液体基质,其中含有蛋白质、核酸和各种代谢物(图1-14)。线粒体内膜有许多蛋白质镶嵌在双分子层脂类系统中,这些蛋白质包括各种运输蛋白、脱氢酶类、电子传递系统和 ATP 合成酶类等,是进行氧化磷酸化的重要部位。所谓氧化磷酸化是指将生物氧化过程中释放出的能量用于合成含高能磷酸键的 ATP。
图 1-14 细胞线粒体的结构
植物的绿色细胞中含有数目不等的叶绿体,它们由两层外被膜所包围, 内含以蛋白质和核酸为基本成分的基质,在基质内有膜系统排列折叠成片层结构,片层间隔扩大成扁平圆盘状胞囊,为类囊体。类囊体彼此垛叠成基粒, 有如叠在一起的硬币,基粒之间由片层连接。这种复杂的片层结构大大增加了膜的面积(图 1-15)。膜上含有可进行光反应的叶绿素等光合色素。由许
多叶绿素分子和蛋白质组装成的捕光复合体就如同天线,它们收集、吸收光能,并将其汇集到反应中心的叶绿素分子上,只有这一小部分位于反应中心的叶绿素分子参加光反应,将光能转变成化学能。叶绿体的基质利用 ATP 的能源和还原辅酶Ⅱ的还原力促使二氧化碳还原成糖,该过程不需光照,故称为暗反应。生物合成与生物氧化正好相反,这是一个利用能量的还原过程。
图 1-15 叶绿体的结构
- 生物膜的信息传递 细胞在其生命活动过程中必须不断从外部获得信息,并作出相应的反应。无论是细胞内外,或是细胞之间的信息传递均有赖于生物膜。信息借助于信号进行传递。从细胞外将信息传递到细胞内的过程称为信号转导。细胞的信号转导系统由受体、信号转导蛋白(也称 G 蛋白) 和效应器三部分组成,它们都位于细胞表面的质膜上。
受体可分为三大类:第一类受体多为数个亚基构成的寡聚蛋白质,是依赖于化学信号分子(如激素等)的离子通道,在细胞间传递信息(称为第一信使)受体亚基接受信号后即改变通道的开关。第二类受体与信号转导蛋白相偶联,它们多为单条多肽链,将信息转给效应器,产生胞内信使(在细胞内传递信息的分子,称为第二信使),从而激活细胞内有关的酶系统。第三类包括一些生长因子的受体,它们本身具有酪氨酸蛋白激酶的活性,可直接激活细胞内的酶系统。
G 蛋白是信号转导系统的重要一环。有许多种 G 蛋白,分别联系不同的受体和效应器,大致可分为刺激性和抑制性两类。因此同样的信号分子对不同的组织细胞可引起不同的效应。效应器可以是一种酶,用以合成或分解第二信使,或者是一种离子通道,以改变膜的离子流和膜电位。膜电位在传导电信号中起重要作用。
值得提出的是,迄今发现的癌基因都是属于信号转导系统蛋白质的编码基因,因发生突变失去控制而成为癌基因。这是因为细胞的正常生长和分裂是受控制的,细胞内外环境条件的改变通过信号转导系统而影响细胞生长和代谢,它的失控将导致细胞恶性生长,即发生癌变。
- 膜的生物合成和人工膜生物膜上含有合成膜脂的各种酶类。脂类的前体物质(如脂肪酸、甘油、含氮化合物等)是在胞液和细胞器基质中合成的, 它们被膜固有的酶利用来合成膜的脂类分子和糖脂。蛋白质是在细胞质中合成的多肽链在合成的一定阶段或合成后被引导并插入生物膜的特定部位。生物膜的合成就是在原有膜上加入新的成分,当膜生长至一定程度后即发生细胞和细胞器的分裂。
利用膜的自组装、自封闭和自分相性质,可以用生物膜的成分重组功能膜,或用脂类分子制造人工膜。通过超声波处理,有机溶剂处理、机械搅拌、高压喷射等方法,可制备出各种大小的人工膜囊泡,称为脂质体。脂质体在理论研究上和实践应用上都有很大价值。
- 细胞器和细胞
细菌和蓝藻都属于原核细胞,这类细胞膜结构比较简单,无细胞器。动植物细胞(包括酵母菌)具有核结构,属于真核细胞。这类细胞内部由双层核膜将原生质分成细胞核和细胞质两大结构与功能区。细胞核是遗传信息贮存场所,在这里进行基因的复制和转录,从而控制着细胞的代谢活动。细胞
核的主要结构有核膜、核仁、染色质及核液等。细胞分裂时染色质凝缩成染色体(图 1-16)。细胞核内合成的 RNA 经加工后由核孔进入细胞质中,在那里进行蛋白质合成。
图 1-16 染色质的各级结构
在细胞质内以膜的分化为基础形成许多重要的细胞器,如内质网、高尔基体、溶酶体、线粒体等。胞液是指细胞质的连续水相部分,大部分中间代谢都在胞液中进行;其间还有各种颗粒、纤维和膜状结构,细胞器即悬浮其中。内质网是分泌蛋白和合成磷脂、糖脂、胆固醇等的场所。高尔基体主要功能是对细胞合成和吸收物质分门别类地进行浓缩、加工和运输,因此细胞的分泌和吞噬都与其有关。溶酶体的膜形成囊泡状结构,是细胞的消化器官。线粒体是重要的分解代谢和能量代谢场所,生物氧化磷酸化在线粒体内膜上进行。植物细胞除上述细胞器外,还有叶绿体、液泡和细胞壁。此外,细胞中还有许多特殊的或细微的结构。图 1-17 为动物细胞的典型结构。
图 1-17 动物细胞的结构
组成生物体的细胞有多种多样,其形状与大小悬殊很大,结构的复杂程度也很不一样。细菌是比较小而简单的细胞,多数细菌细胞直径为 1~2 微米,呈球状或杆状。是否有比细菌更小的细胞?目前发现的最小与最简单的细胞是枝原体,它们中很多是人与动物的病原体,其直径只有 0.1~0.15 微米,只有细菌体积的千分之一。为维持细胞独立生活,估计至少要有 100 个左右基因,以便编码几十种维持细胞最基本代谢过程所需要的酶。这样看来, 细胞体积的大小在理论上是有下限的,枝原体已接近此下限。比枝原体更小的病毒是不能离开细胞独立生活的,它们应看成是具有感染能力的细胞成分。动植物细胞比较大,结构也较复杂,其直径多数在 15~30 微米之间。也有不少例外,例如鸟卵细胞的直径可达数厘米,这是由于卵中含有丰富的营养物质,以供受精后发育的需要。
在种类繁多的细胞世界中,根据其进化方向与结构特征,可以分为原核细胞与真核细胞两大类。一般认为原核细胞与真核细胞起源于共同的原始细胞,大概从 30 亿年前这两类细胞就开始分道扬镳了,它们不仅在结构复杂程度上有很大差别,而且是沿着两条不同的演化途径发展。以细菌为代表的原核细胞主要特点为:结构小巧、表达高效、分化较低、生长速度快。动植物等真核细胞的特点为:结构复杂、调节精确、功能分化、适应潜力大。原核细胞以快速生长见胜,其结构和代谢无不均表现出这一优势。剩菜剩饭隔一夜就变馊了,就因为长满了细菌的缘故。真核细胞具有庞大的基因组,衍生出多种多样的复杂结构和代谢类型,以各种不同的方式去适应和征服自然。世界上现存的动植物物种大约有二百万种以上,我们所见到的绚丽多彩的生物界主要就是由真核细胞组成的。
从细胞形态结构上来看,原核细胞的膜结构并不发达,只有质膜和由质膜凹陷形成的中膜体,不形成核和其他细胞器;真核细胞的膜结构很发达, 形成核和其他细胞器。核结构的出现使转录(RNA 的合成)和翻译(蛋白质的合成)在空间上和时间上都被分隔开来,基因表达的调控得以在不同水平
上更精确地进行;而在原核细胞中转录和翻译往往是同时进行的。从染色质的结构上来看,原核细胞的 DNA 仅局部覆盖以蛋白质,并不形成核小体和染色体的高级结构;真核细胞的染色质存在多层次的高级结构。染色质的组装结构不同,决定了它们复制和分配方式的差异。原核细胞的繁殖以直接分裂为主,真核细胞则进行有丝分裂。从基因组的结构来看,原核细胞的基因组小而紧凑,往往在功能上彼此有关的基因组成一个转录和调控的单位,称为操纵子,它们由同一控制序列所启动。真核细胞的基因组庞大而复杂,有较大比例的调控序列和重复序列,编码蛋白质的基因不组成操纵子。原核细胞和真核细胞的基因结构也有较大差别,前者是连续的,后者是不连续的,编码序列往往被一些非编码的居间序列所分隔开。因此真核细胞转录的 RNA 要经过拼接,才成为有功能的成熟 RNA。由此可见,原核细胞和真核细胞主要区别在于两者遗传物质的组织结构方面,即两者在核、染色质、基因组和基因结构上有明显的不同。了解这一点很重要,在进行基因操作时必须考虑到各自的结构特点。
- 多细胞生物的组织和器官
真核生物除原生动物、单细胞藻类和酵母菌外,多数为多细胞生物。在多细胞生物中,各类细胞有不同程度的分化。所谓细胞分化是由于生物在发育过程中永久性地关闭了细胞基因组的某些基因,因而使它们表达的性状不同。例如,动物肝脏细胞和肌肉细胞的基因组是一样的,但是它们表达的基因不同。肝脏细胞经分裂后产生的子代细胞仍然是肝脏细胞,因为肝脏细胞不需要的基因已经被关闭了。细胞分化是在染色质结构水平上控制的。基因组某些区域发生异染色质化,即这部分基因被组装成高度压缩的异染色质, 从而不再能表达。这种异染色质化具有“记忆”的能力,就是说子代细胞能保持该区域处于异染色质化。癌组织失去了分化控制,因而又恢复了胚胎时期的旺盛生长。研究细胞分化和再分化的规律,无论在理论上和实践上都有十分重大的意义。
细胞经分化形成不同的类型,分别集合成群,称为组织。由各种组织构成执行一定功能的器官。由许多器官联合组成的一套结构称为系统。例如, 高等植物的组织可分为表皮组织、维管组织和基本组织三大类,当然它们还能进一步细分。由这些组织构成根、茎、叶三种器官,它们均属于营养系统。花、果实、种子三种器官则属于生殖系统。又如,高等动物组织可分为上皮组织、结缔组织、肌肉组织和神经组织四大类,由这些组织分别构成心、肝、肺、胃、肠、肾、眼、鼻、耳、脑等器官,这些器官分属于循环系统、消化系统、呼吸系统、排泄系统、神经系统等。由此构成生物个体不同层次的结构,这些结构都是在生物大分子的基础上与其他有机和无机分子及水逐级装配而成的。
